This grew out of a talk I had the opportunity to give to Novella Carpenter’s Urban Agriculture class at University of San Francisco in 2026.


I was inspired for a title by this article authored by Dani Zamir, a big deal in the world of plant breeding, real plant breeding, not just amateur breeding like me.
But even at my level, I see it as an art and a science. It is an art because it is a creative expression, I am guided by intuition and joy and curiosity and beauty.
Which leads to the science: taking careful notes, trying specific experiments to see what will happen, learning more to be able to guess what might happen and then compare the actual results.
I am inspired in my artistry by my mom, natural perfumer and alchemist Mandy Aftel, by Gregor Mendel, by Charles Darwin, by Charles Ricks, Tom Wagner, Craig LeHoullier, Fred Hempel, Karen Olivier, Bill Yoder, and the so many others working to make beautiful, tasty, healthful, adaptive tomatoes.
In trying new varieties, I estimate that 1 in 20 will be unusually interesting, vigorous and flavorful. So I try many. The same for creating new varieties from crosses, I am estimating that 1 in 20 more or less might be interesting, and I am not attached to any particular idea. I am viewing it as creative work, as searching and sifting. Like alchemy, I am starting with the best I can find and then trying to make something even better. (Better to my tastes, and taste is subjective).

These are some of the tomatoes you can grow today. Besides the visual beauty, there is a kind of beauty in a plant that grows well, and in food that tastes fantastic. So to be able to have all three is a really beautiful thing.
Saving seeds can help select for vigor and flavor within the local environment; if you save and keep seeds year after year, this is like the process of heirloom seed saving. It’s wonderful to grow your favorites year after year, and to share them with friends and family and even pass them down generation after generation.
Cross-breeding allows new combinations of colors and shapes and flavors, and the possibility to incorporate the genes of tomato wild relatives into them for new traits like disease resistance, productivity, flavor and new color combinations.
(I created this collage mostly with my own photos, and borrowed a few elements from MerakiSeeds and TomatoFifou).

Saving seeds is the first step.
You can save seeds from existing varieties, year after year selecting the ones that grow most vigorously and taste best to you. That’s very fulfilling in itself.
If you’re saving a few, all you need is a paper towel, a pen and a piece of tape.
If saving more, then cups or buckets, and a strainer, and paper plates (for drying); ferment 3-5 days and pour off scum and add water and swirl pouring off bad floating seeds and rinse until clean and strain, spread thin on paper plates and dry for a few weeks.
Hybrid varieties (F1) seeds won’t be “true” (the same as the parent) and sometimes have unforeseen traits like male-sterile (in which you would need to hand-pollinate each and every flower to get fruit) and so can be frustrating to get involved with, but also an experiment. Some varieties are patented and the seeds are not legal to save.

It’s not necessary, but if there’s one thing that makes a big difference, it’s saving your seeds in the fridge. My germination rates are really excellent. I have ten year old seed that are almost all germinating. For fresh seed, make sure they air dry for a few days, before putting them in a paper envelope (breathable) and store in gallon bag or Tupperware with desiccant packs. They like to be cold and DRY. This is worth doing if you are serious about any kind of seed saving.
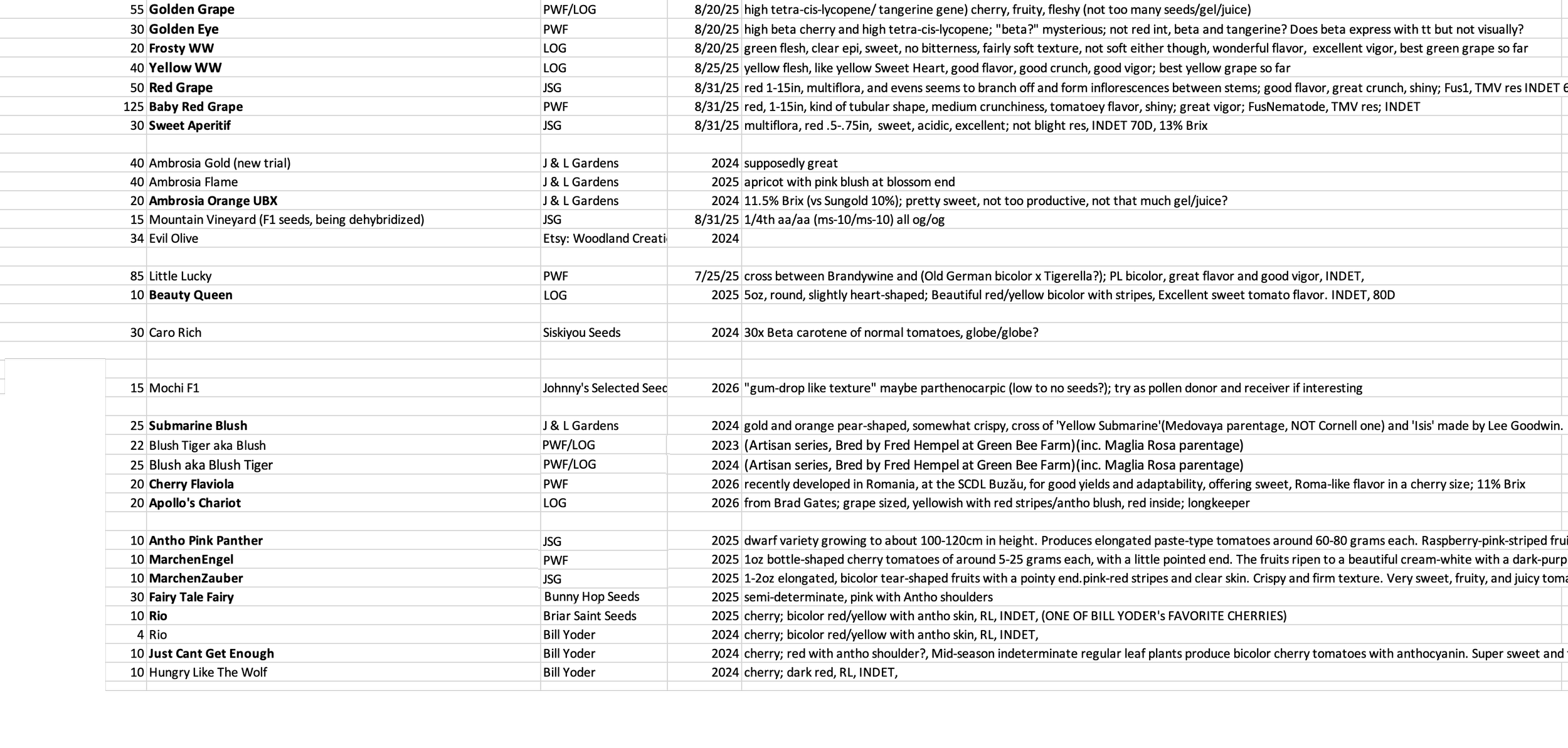
Organization is important. I use a spreadsheet to count number of seeds, origin, date, and notes on genetics, flavor, etc.

To start my seeds, I use a well-draining coir based soil mix, full spectrum grow lights, and heat mats at 80 degrees. This is after 1 week, I went on to pot them up into 4 inch pots between 1 and 2 weeks.

After 1 month, these are ready to be set out and put in even bigger pots or planted in the ground after about a week or two of adjusting and growing outside.
(The nighttime temp must be 50 degrees or above to avoid potential damage to tomato plants, so don’t start seeds more than a month before that if you’re planning on setting them outside then).
I started them here in early-March, and set them out mid-April (partially covered as they adjust to sun exposure).

Once you know how to save seeds, you can also try cross-breeding different varieties, and save seeds from those to end up with new varieties.
It typically takes about seven cycles of saving seeds and growing plants and saving seeds and selecting for certain traits to stabilize a new variety.
This can mean 4-7 years. Adapting to them to your local environment and to your specific likes is a really neat process, and watching something come
into being before your eyes (over several years) is a real pleasure in itself and sometimes a test of patience.
You need to know something about the tomato biology and the flower parts in order to make a cross.
Tomato flowers are known as “perfect” flowers, containing male and female parts.
The male parts: stamen including the anthers and filaments- on a tomato the anthers are fused into an anther cone.
The female parts: carpel containing ovary (and ovules inside) with style and stigma on end (pollen-receiving part).
Tomato flowers are able to self-pollinate and usually do- more than 95% of the time, especially because the modern varieties have a stigma that is not exserted, does not protrude out of the flower.
The anther cones mature and dry out or “dehisce” and release the pollen before the flower is fully open, fertilizing the ovules.
In order to make a cross, the anther cone must be removed just after the sepals (outer green things covering the petals) begin to lift open, but before the petals have begun to open and the pollen has been released.


The flower on the left is opened, and too late to use as the mother in a cross (because it’s already self-pollinated), but is right to collect pollen from.
In the inflorescence on the right, there is one flower in which the sepals have just begun to open, and is a prime candidate for crossing.


On the left, I’m about to dab the stigma of a flower I’ve emasculated (removed the anther cone from) into some of the pollen I’ve vibrated into a little container using a cheap electric toothbrush.
Pollen lasts at least a week under normal conditions, and up to a month dried and stored in the fridge.


These are good signs.
Once a fruit is well on its way to forming, I put a teabag around it to keep track of it and keep it safe.


These are bad signs, not successful pollinations.
I try pollinating multiple times over a few days, and stop when I see the good signs (or bad signs obviously).
I use a twist tie with the names on it until a fruit forms, then cover it with a tea bag.


When I started it was in 3 or 5 gallon pots. Make sure to water when dry!
Here are several successful crosses collected and ready to be saved.

Seeds to add to the possibilities of what to grow next year.

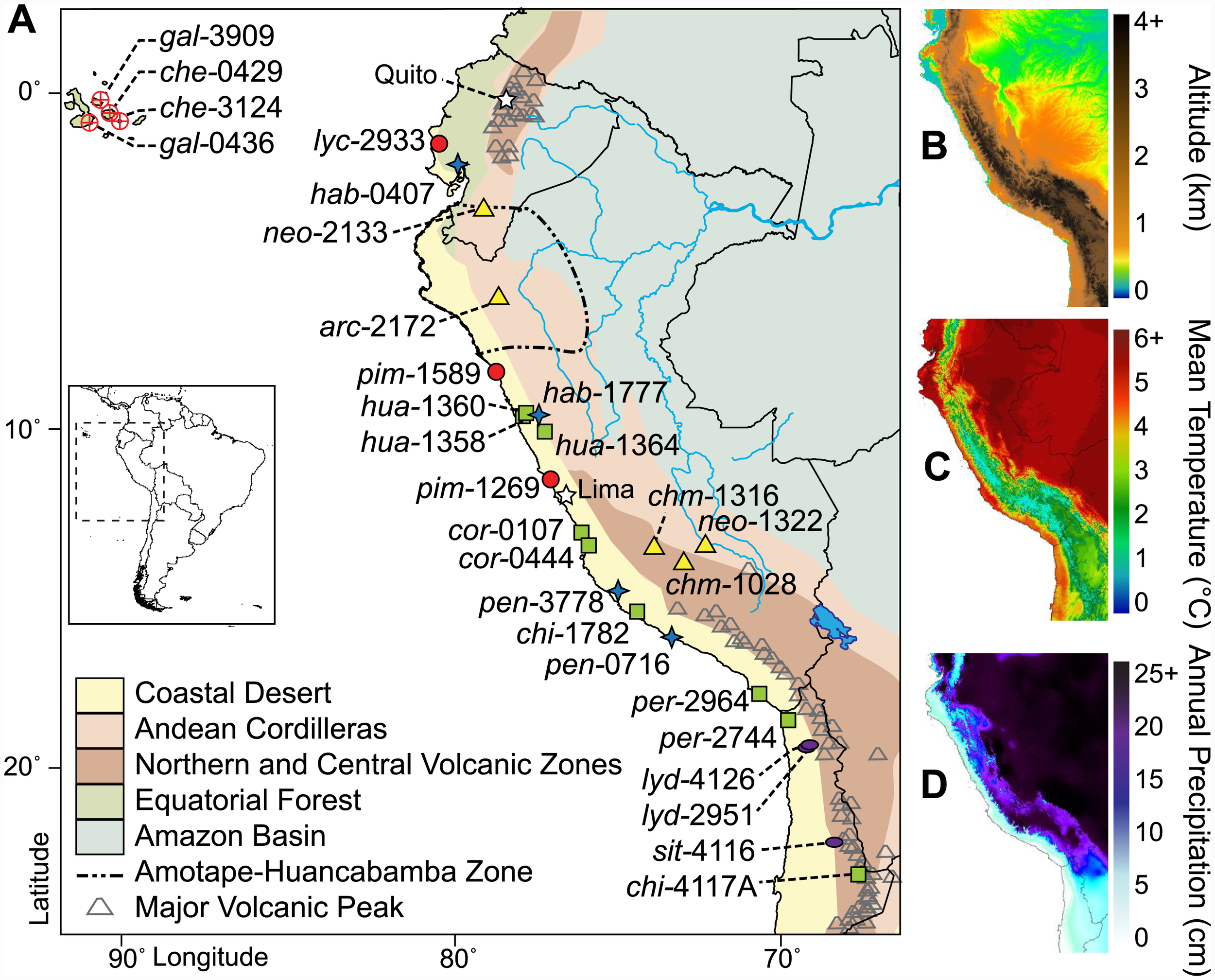
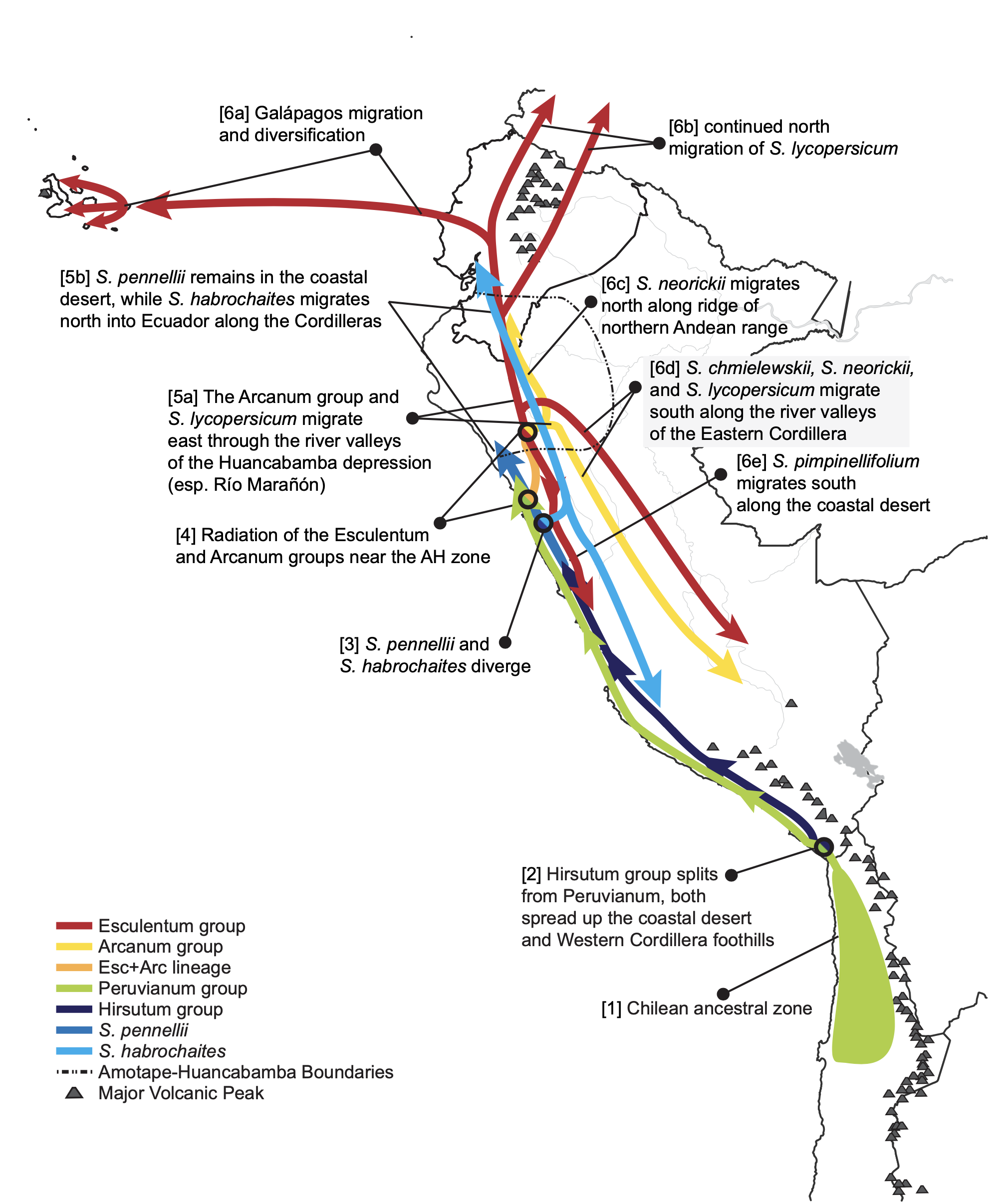
This is where tomatoes came from, and there are still wild relatives living there to this day. (the little shapes on the map)
For any plant it is useful to know where it came from, it’s original environment, the conditions it evolved in, adapted to; the temperature range, sun exposure and amount precipitation. Tomatoes originated in Peru and Ecuador over 80,000 years ago, where it is warm and relatively dry.
The genes evolved from that place, from those conditions; an interplay between the plants and their environment, and so that is where a discussion of the genes begins.
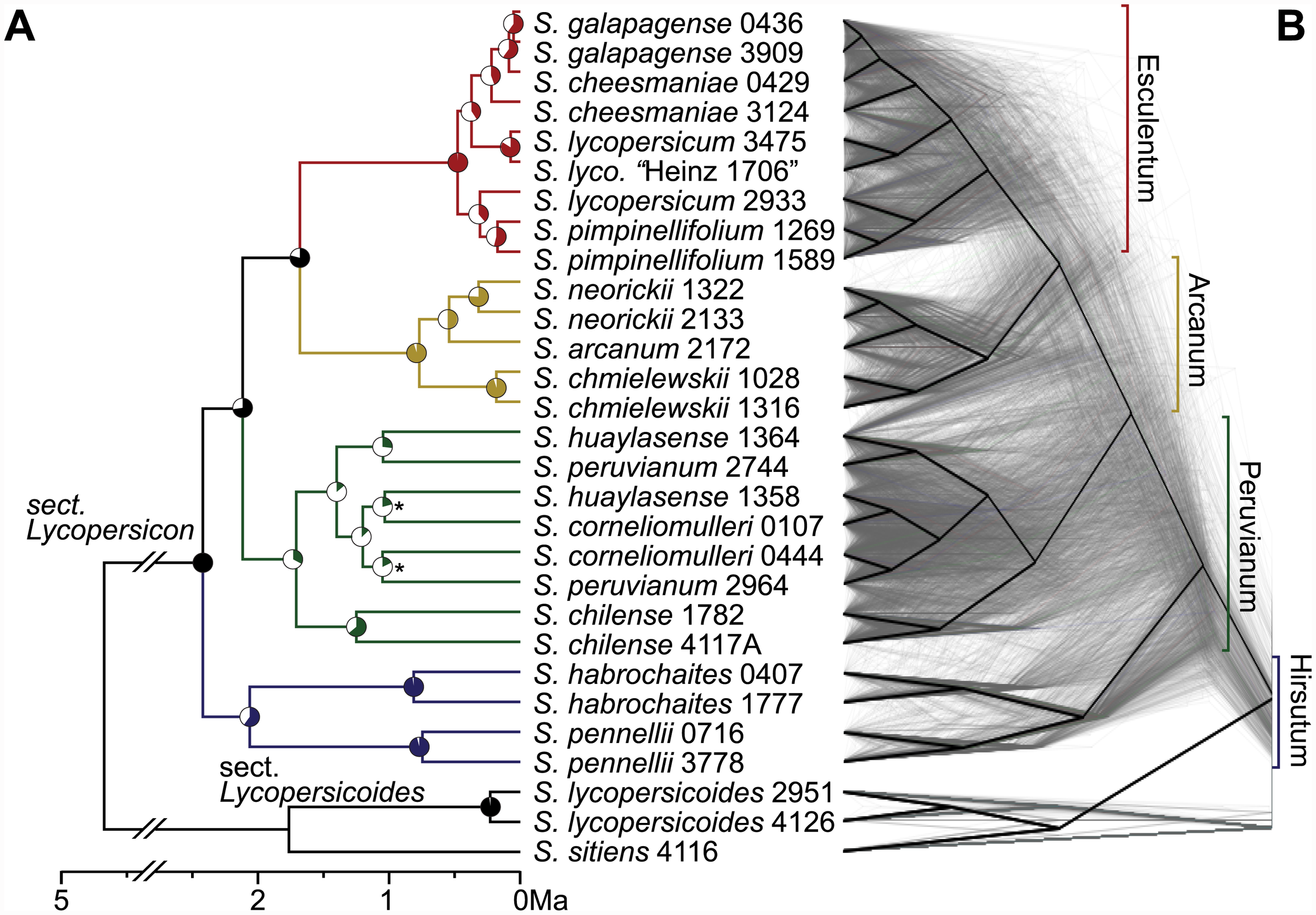

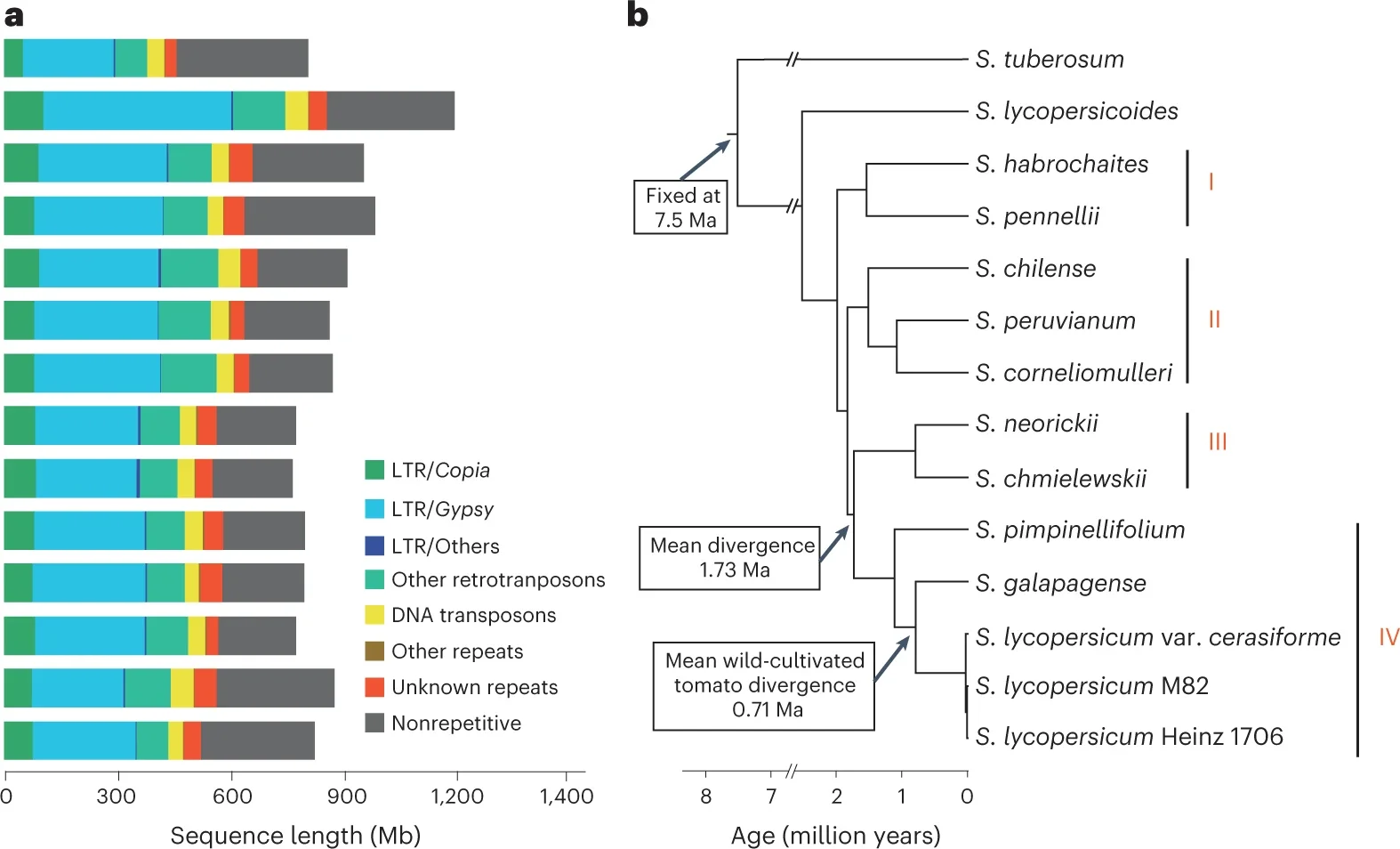
As these research papers show, tomatoes are related to other species of Solanums, some more closely related and able to interbreed with tomatoes. The closest related is called Solanum pimpinellifolium (Currant tomato) which phylogenetic studies show is the common ancestor of all cultivated tomatoes.
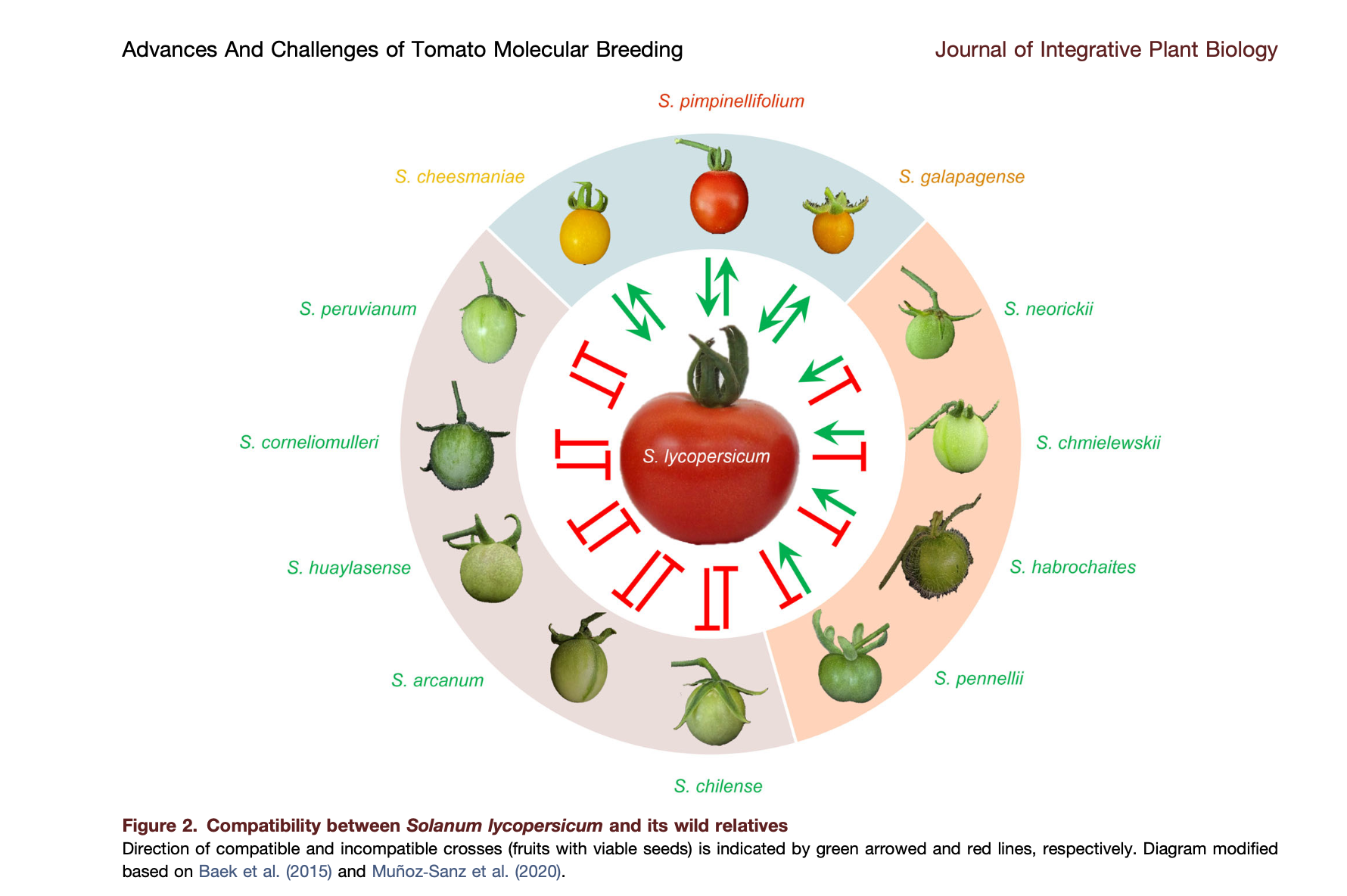
This diagram illustrates what the fruit of many of those wild relatives look like, and indicates whether that species is able to interbreed with the tomato (Solanum lycopersicum).

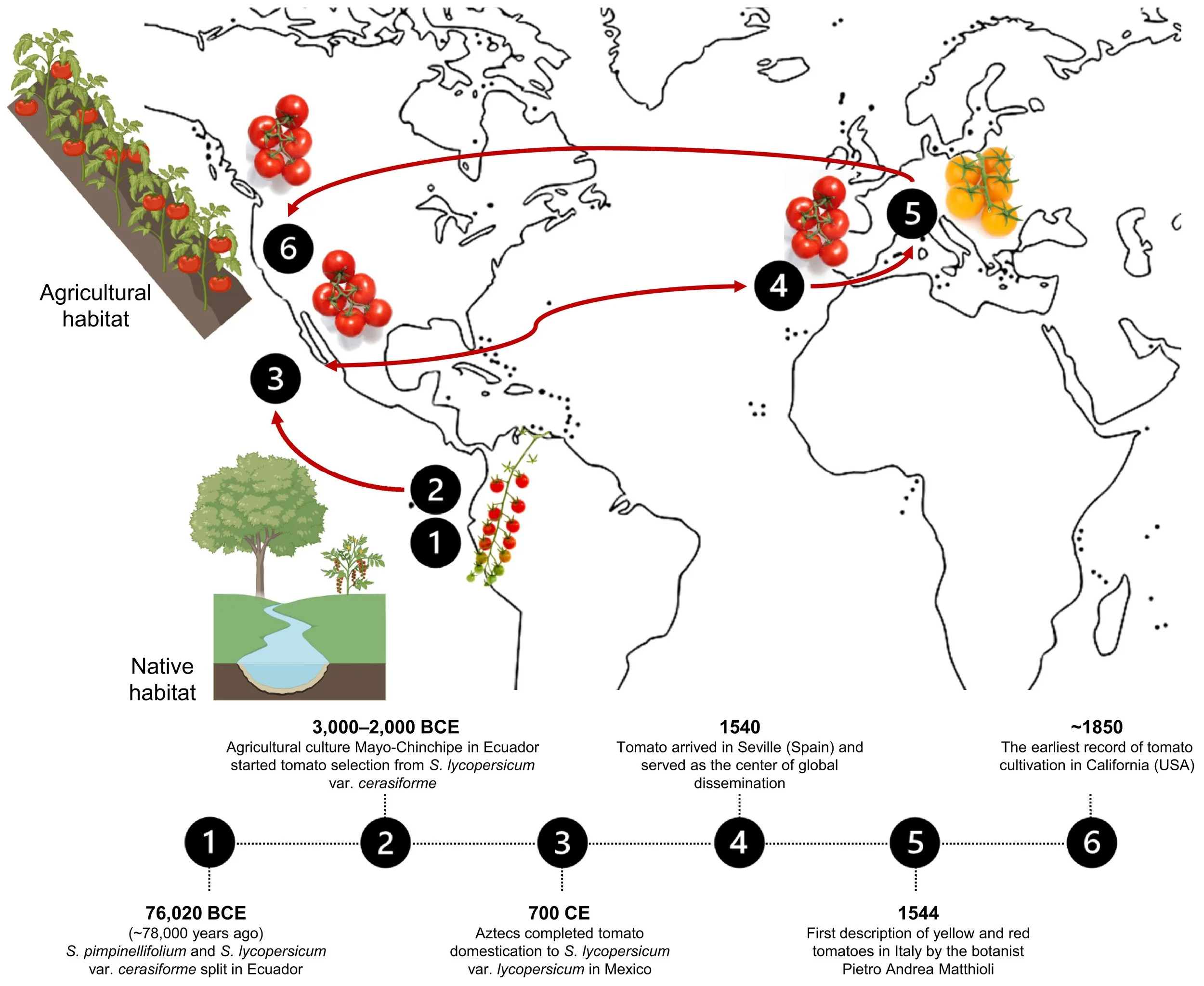

This paper shows their journey of domestication.


This image illustrates some of the variations that tomatoes picked up during their domestication.
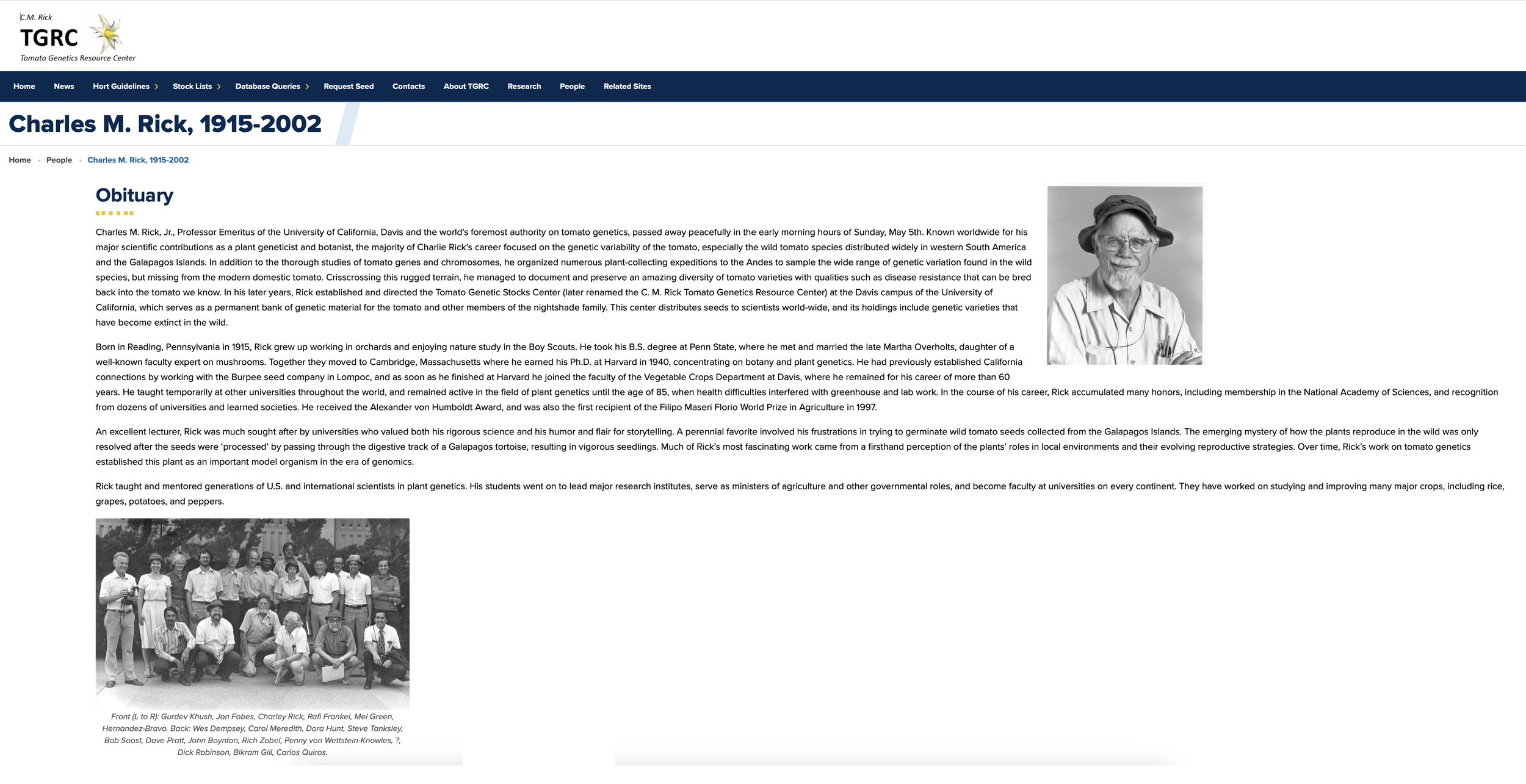
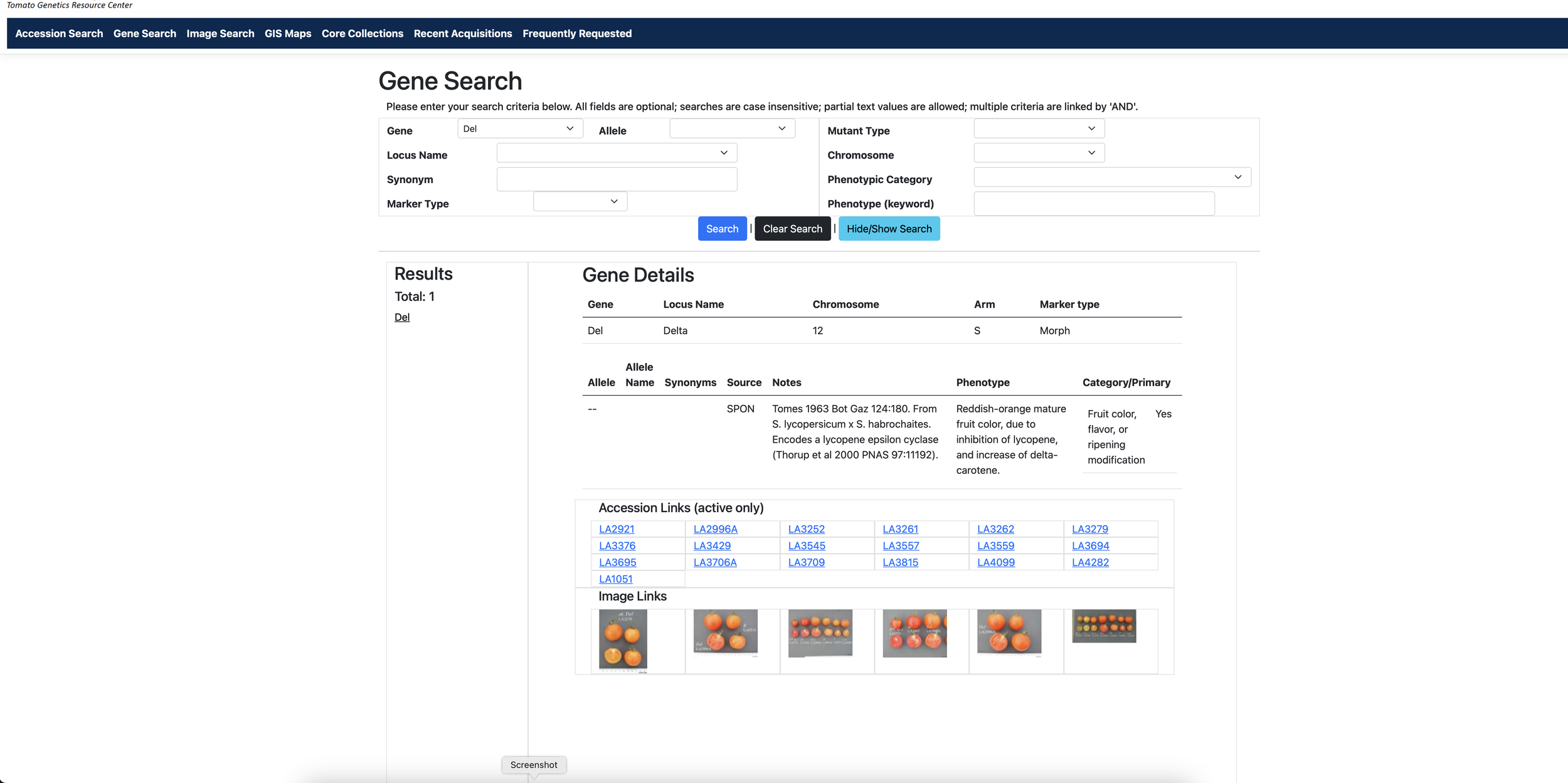
Charles Rick really opened the door to incorporating wild species genes into tomatoes and to understanding tomato genetics. He went out and collected those wild relatives and helped figure out how to cross-breed their genes into tomatoes. His collection is maintained as a resource for tomato breeding to this day at the UC Davis C.M. Rick Tomato Genetics Resource Center.
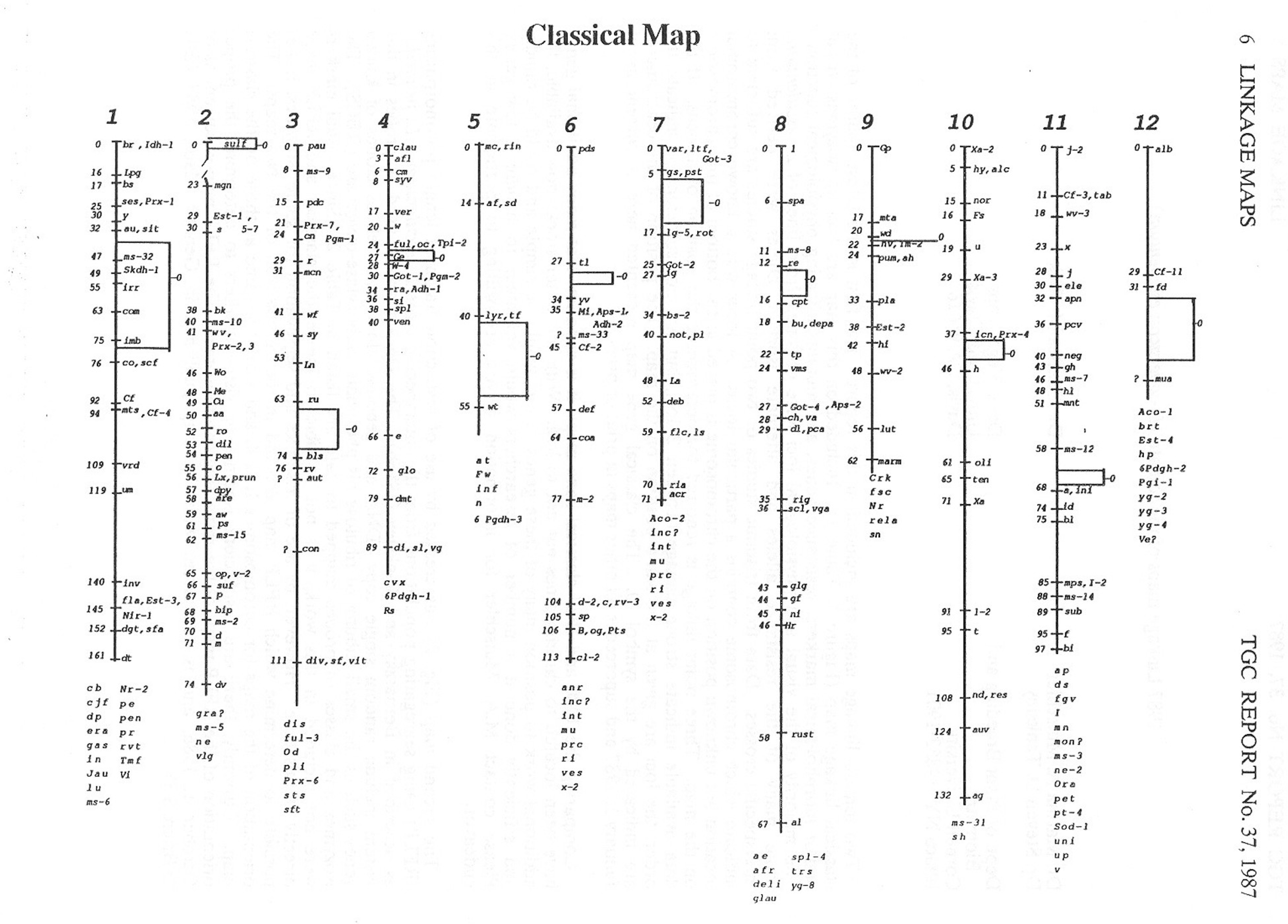
The tomato contains 12 chromosomes, and thanks to the pioneering work of Charles Rick and so many others, the location of many genes are well-documented (some locations have been refined since; from TGC Report No. 37, 1987)
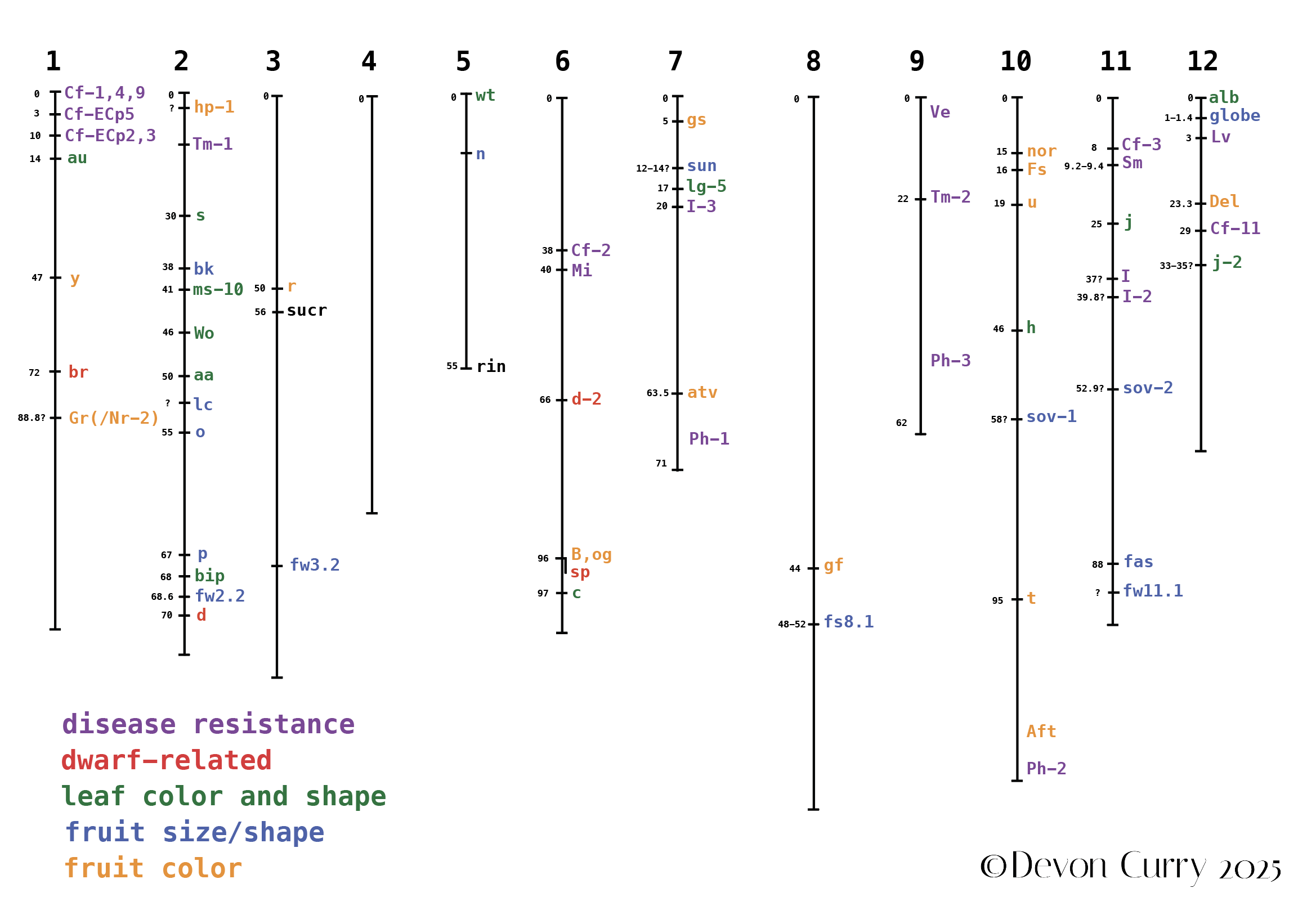
This is a working copy I use to keep track of genes that are relevant to me and to look for potentially linked genes,
so that I can better interpret results of my breeding experiments.

Color in tomatoes is like a layering, starting from the inside out and from the top of the carotenoid creation process to the bottom:
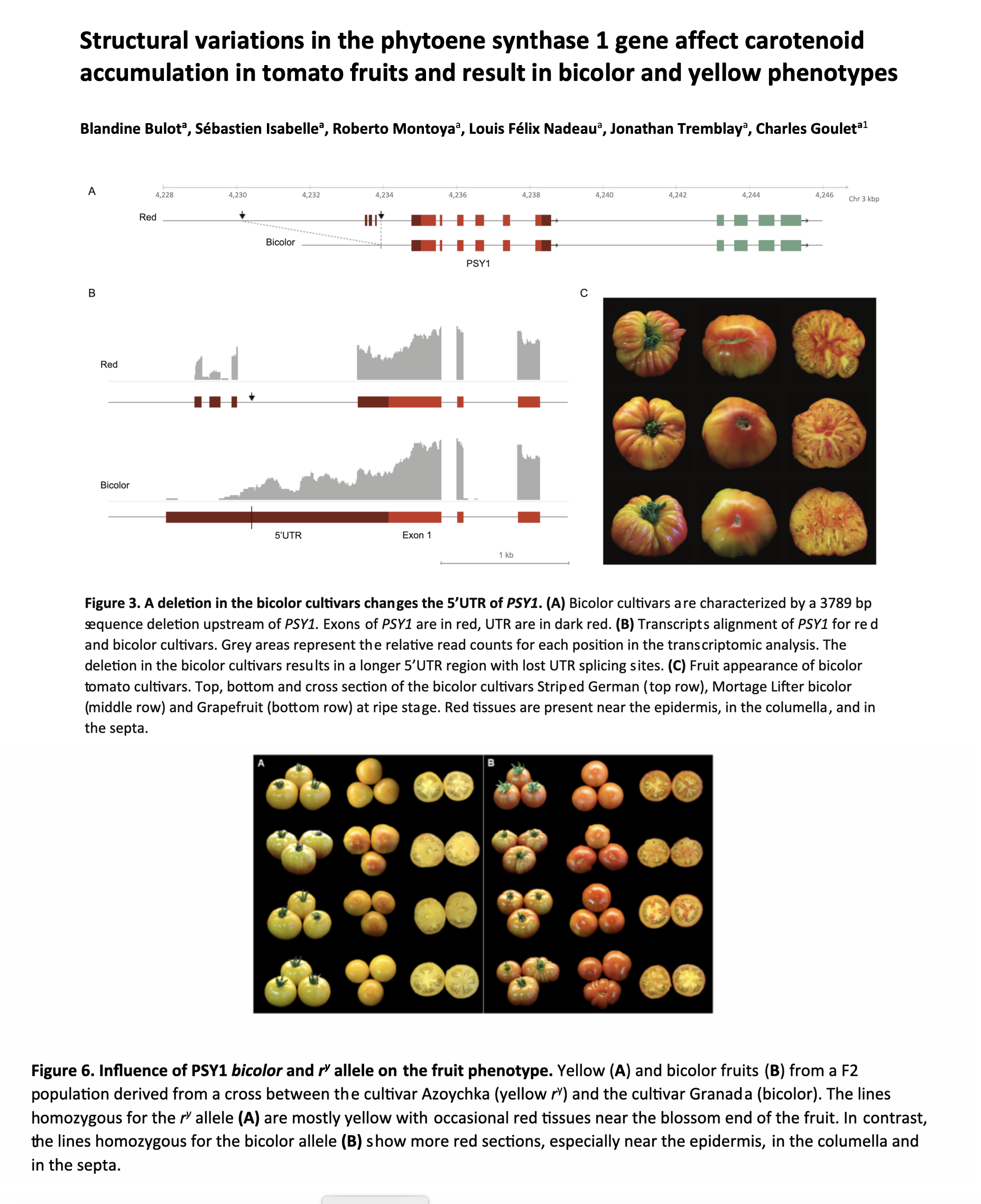
At the R locus, you can have the wild-type (R) red allele, the mutant (r) yellow allele or a mutant (ry or bicolor) bicolor allele.
What’s happening is that that a enzyme called phytoene synthase 1 (PSY1) is partially (bicolor) or totally (yellow-flesh) dysfunctional.
PSY1 is a precursor to all carotenoid development, so a tomato with two copies of the (r) allele that creates no PSY1 has something like 80 percent lower carotenoids than a tomato with the (R) allele.
In the bicolor allele, there is a very ineffective copy of the gene; it has a huge insertion it it’s promoter, so it takes longer to read the gene, and makes only small amounts of what becomes lycopene. That lycopene then naturally goes to certain places in the fruit first as it develops, and thus the pattern of red streaks.
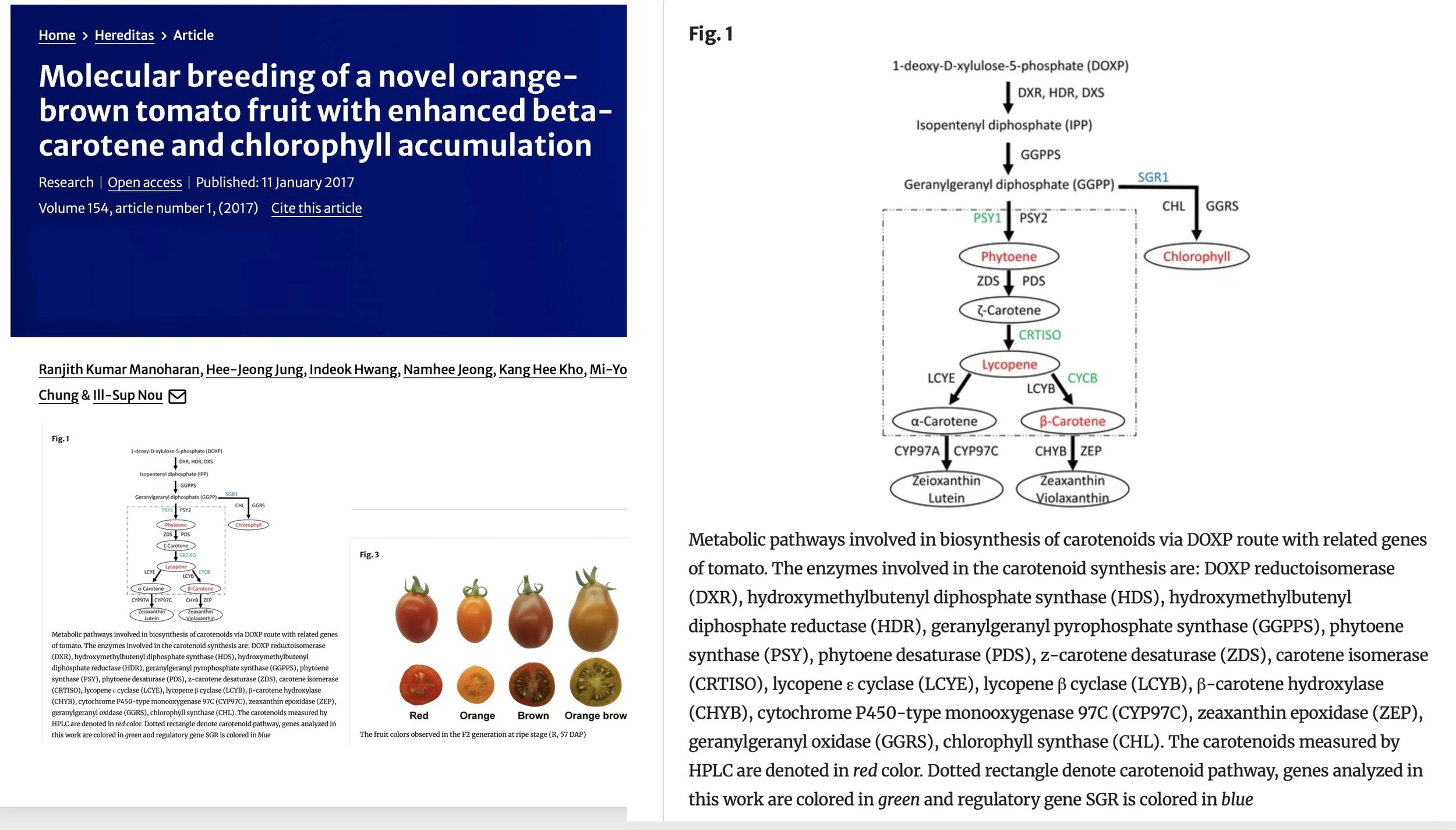
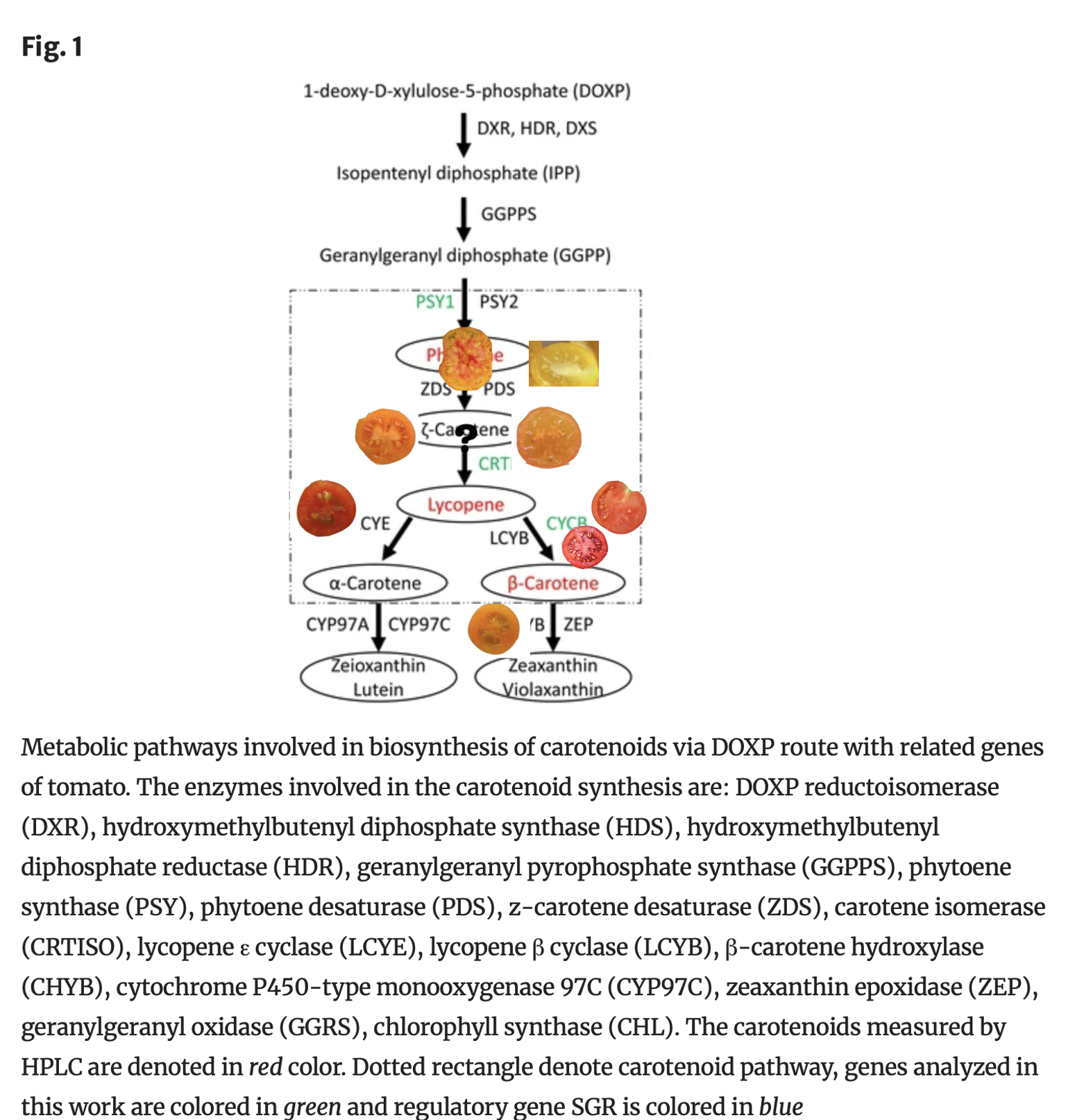
The next step in the carotenoid pathway is called carotenoid isomerase (CRTISO) created by the T gene.
This gene allows orange-colored prolycopene (aka tetra-cis-lycopene) to be converted into red-colored lycopene.
If the gene is dysfunctional as in the mutant (t) allele, then the tomato accumulates prolycopene, and no lycopene is made; it is orange, “tangerine orange” after a representative tomato which has that gene. Typically tastes fruitier than beta orange and is solid orange.
If there is at least one copy of the T allele to create CRTISO, then red lycopene is made. This is the typical red tomato.
But from there, if there is a copy of the Beta gene as well, the lycopene that is created is then converted into orange beta-carotene, this is a “beta orange,” typically tastes more carroty, and often has reddish center.
Or if there are two copies of the recessive B og allele (null-beta “old-gold”) then no beta-carotene at all is made, which results in more lycopene remaining. These are known as old-gold or “crimson” tomatoes, which are a redder color of red, and have red locules (gel surrounding seed is red instead of green). (They are called old-gold because of their characteristic flower color pattern).
Or if the Del gene is present, then the orange-colored Delta-carotene is created, converted from lycopene. This is a more unusual gene.
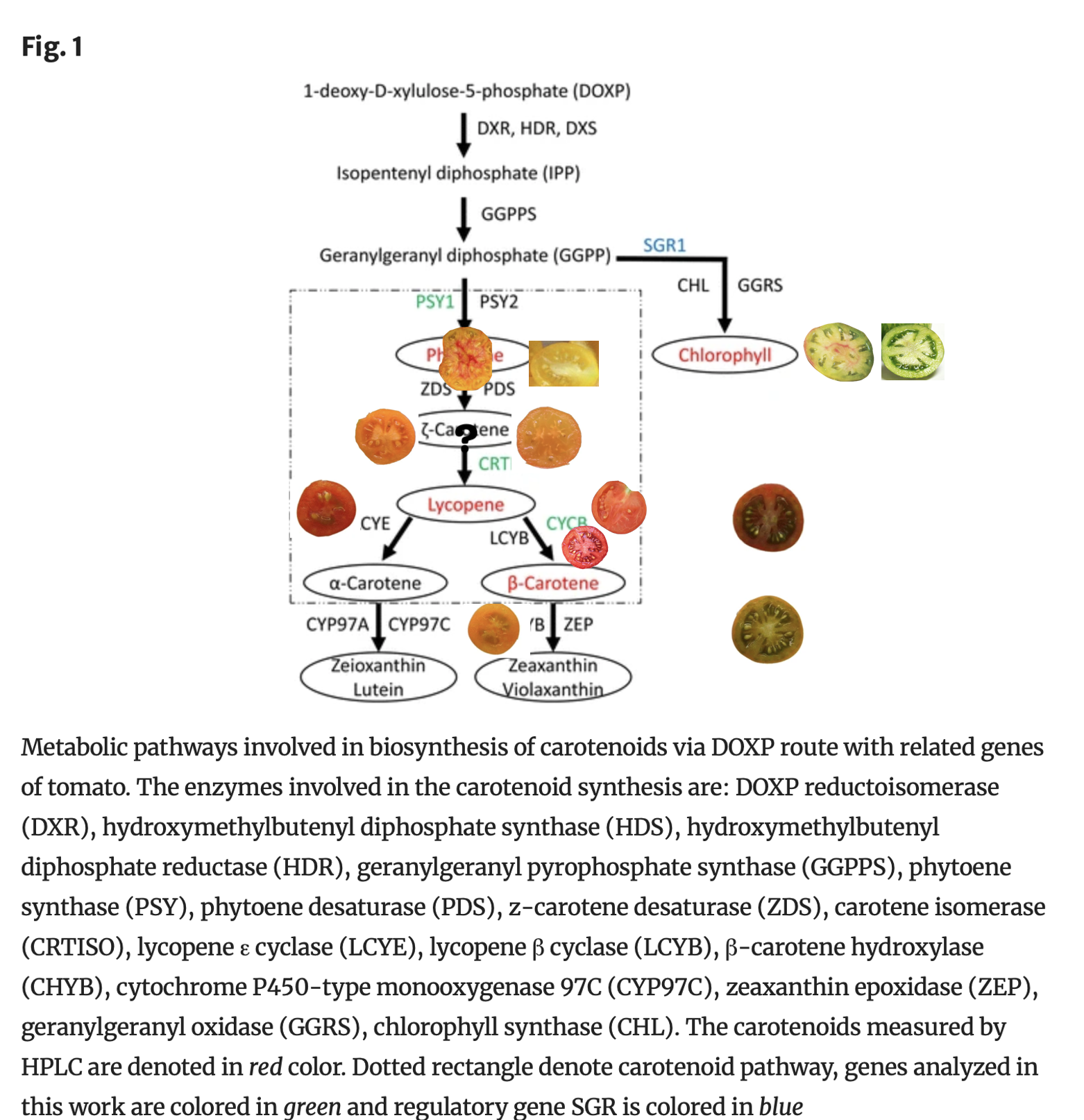
The next layer is when the (gf)greenflesh gene is present. This combines with the carotenoid genes to make different colors.
With an otherwise red tomato, the greenflesh gene would make it a brown tomato
•For a pink tomato (red with clear epidermis (y gene, next topic), the greenflesh gene would make it a purple tomato
For orange , it would become ochre (brownish green color)
For yellow, it would become yellow green, “green”
•For white/ivory (yellow with clear epidermis (y gene, next topic), it would become “clear green”
With a bicolor tomato that’s yellow with red in the center, it would become green with red in the center

The next layer to understanding tomato color is at the Y locus:
There can be the wild-type (Y) yellow epidermis (aka skin) or the mutant (y) clear epidermis.
This accounts for the difference between:
Red(yellow skin), brown(yellow skin), green(yellow skin), yellow(yellow skin), orange(yellow skin) and
pink(clear skin), purple(clear skin), clear green(clear skin), white/ivory(clear skin), light orange(clear skin)
Two more layers that can occur in combination with any other combination are the Aft (anthocyanin fruit) gene, an introgression from the wild species Solanum chilense that causes the purplish or “blue” color on the skin, and the greenstripe (gs) gene, which can be other colors besides just green.
And there’s also a Fruitstripe (Fs) allele that provides a different pattern of stripes, radial ones that meet at the bottom.
Research papers (in order of appearance)
Improving plant breeding with exotic genetic libraries*
https://www.nature.com/articles/35103590
Phylogenomics Reveals Three Sources of Adaptive Variation during a Rapid Radiation
https://journals.plos.org/plosbiology/article?id=10.1371/journal.pbio.1002379
Super-pangenome analyses highlight genomic diversity and structural variation across wild and cultivated tomato species
https://www.nature.com/articles/s41588-023-01340-y
Molecular breeding of tomato: Advances and challenges
https://onlinelibrary.wiley.com/doi/full/10.1111%2Fjipb.13879
The Tomato's Tale: Exploring Taxonomy, Biogeography, Domestication, and Microbiome for Enhanced Resilience
https://apsjournals.apsnet.org/doi/10.1094/PBIOMES-09-23-0091-MF
Cultivated Tomato (Solanum lycopersicum L.) Suffered a Severe Cytoplasmic Bottleneck during Domestication
https://www.mdpi.com/2223-7747/9/11/1443
Genomic Evidence for Complex Domestication History of the Cultivated Tomato in Latin America
https://academic.oup.com/mbe/article/37/4/1118/5679792
Charles Rick Obituary at TGRC
https://tgrc.ucdavis.edu/people/charles-rick
TGRC Gene Search Page
https://tgrc-mvc.plantsciences.ucdavis.edu/Genes/Search
Genetic basis of fruit quality traits in processing tomatoes*
https://www.sciencedirect.com/science/article/pii/S2666154325004673
Distribution of SUN, OVATE, LC, and FAS in the Tomato Germplasm and the Relationship to Fruit Shape Diversity*
https://academic.oup.com/plphys/article/156/1/275/6111492
Development and validation of the OVATE gene-based functional marker to assist fruit shape selection in tomato
https://pmc.ncbi.nlm.nih.gov/articles/PMC8546015/
Molecular and genetic regulations of fleshy fruit shape and lessons from Arabidopsis and rice
https://academic.oup.com/hr/article/10/7/uhad108/7191294
Identification and characterization of GLOBE, a major gene controlling fruit shape and impacting fruit size and marketability…
https://vanderknaaplab.uga.edu/files/2024/05/Sierra-et-al-2021.pdf
Structural variations in the phytoene synthase 1 gene affect carotenoid accumulation … result in bicolor and yellow phenotypes*
https://www.biorxiv.org/content/10.1101/2022.10.29.514357v1.full
Epistasis in tomato color mutations involves regulation of phytoene synthase 1 expression by cis-carotenoids
https://www.pnas.org/doi/10.1073/pnas.1214808109
Molecular breeding of a novel orange-brown tomato fruit with enhanced beta-carotene and chlorophyll accumulation*
https://pmc.ncbi.nlm.nih.gov/articles/PMC5226094/
Characterization of Tomatoes Expressing Anthocyanin in the Fruit
https://journals.ashs.org/view/journals/jashs/133/2/article-p262.xml